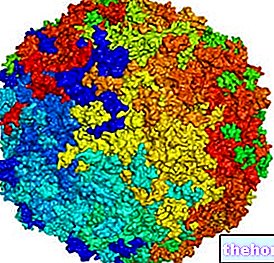
Вируси се могу умножавати само унутар ћелије домаћина, експлоатишући њен метаболички апарат и користећи сопствене генетске информације; међутим, до размножавања долази само у ћелијама подложним вирусу, односно опремљеним специфичним површинским рецепторима и способним да изведу репликативне фазе његовог генома.
Мултипликативни процес је подељен у различите фазе:
1. фаза: напад или адсорпција вируса на ћелијску мембрану;
2. фаза: продирање вируса или његове нуклеинске киселине у цитоплазму ћелије;
3. фаза: свлачење или помрачење (губитак вирусних омотача и излагање нуклеинске киселине);
4. фаза: репликација (синтеза макромолекула, односно ДНК, РНК и вирусних протеина); вируси имају различите стратегије репликације и сваки се размножава на другачији начин, експлоатишући ензиме и органеле ћелије домаћина;
5. фаза: састављање (формирање унутар ћелије - у језгру или у цитосолу - капсиде; унутар ове овојнице убацује се вирусна ДНК, формирајући нуклеокапсид);
6. фаза: ослобађање или бијег вируса из ћелије.
Уз неке изузетке, циклус репликације вируса је веома брз и завршава се за 8-24 сата. Свака од ових фаза је сложена и типична за сваку врсту; у ствари, постоји велики број репликативних стратегија и механизама; прве две (адсорпција и пенетрација) и последња (излаз), на пример, разликују се у зависности од тога да ли вирус има перикапсид или не. Док бактериофаги убризгавају своје нуклеинске киселине директно у цитоплазму ћелије домаћина, те животиње улазе путем пиноцитозе и ослобађају се лизом ћелије и пиноцитозом; током овог проласка нови вириони добијају фосфолипидни омотач и након напуштања могу инфицирати нове ћелије.
Напад, продор и репликација вируса
Голи вируси улазе у ћелију микропиноцитозом, која се назива и виропепса, то јест са истим биолошким механизмом који користи за интернализацију корпускуларних супстанци испод 1 μм. Једном у цитоплазми, ћелијске протеазе варе капсид, а нуклеинска киселина (вирусна ДНК) се ослобађа у цитоплазму.
Напад вируса на ћелију посредују протеини звани антирецептори, присутни на вирусном капсиду и на вирусном перикапсиду, који препознају молекуле или протеине присутне на површини ћелије и називају се рецептори. Стога је фаза адсорпције посредована интеракцијом између антирецептор и рецептор.
АДСОРПЦИЈА: стереохемијска интеракција између специфичних хемијских група изложених на спољној површини осетљиве ћелије (рецептори) и вириона (антирецептори).
ХИВ, на пример, углавном напада Т-помоћне лимфоците, јер има антирецепторе који препознају специфичне протеине изложене на њиховој ћелијској површини.Антирецептор вируса ХИВ-а је перицапсидни гликопротеин, назван ГП120, док се онај Т лимфоцита назива ЦД-4 ; из тог разлога је Т -помоћни лимфоцит познат и као Т4. Након везивања, вирус може ући у ћелију на два начина:
фузија споља: перикапсид се спаја са ћелијском мембраном и испушта у цитоплазму (типично за ХИВ и обложене вирусе);
фузија изнутра: вирус улази у везикулу пиноцитозом. Једном у цитоплазми, перицапсид се спаја са мембраном везикула и капсид се ослобађа у цитоплазму, као што се то дешава на пример са вирусом грипа и уопште са голим.
Очекивано, постоје многе варијације у начину на који вирус улази у ћелију домаћина.
РЕПЛИКАЦИЈА: вируси имају различите стратегије репликације, условљене типом нуклеинске киселине садржане у капсиду; током репликације, уопште, вируси производе две врсте протеина: ране (ензимске и регулаторне природе, попут полимераза) и касне (структурне, које ће формирати капсид и перицапсид). У сваком случају, репликација подразумева пре свега „промену и преусмеравање метаболизма домаћина“, што омогућава вирусу да умножи сопствени геном.
Последња фаза је излазак нових вируса из ћелије (који следи након састављања капсиде у језгру или у цитоплазми). Уопштено говорећи, голи вируси излазе лизом ћелије; међутим, код оних обложених, неки вирусни протеини током репликације, одговорни за формирање перикапсида, идите да се уметнете на једну од мембрана ћелије домаћина (на пример цитоплазматску мембрану, нуклеарну или Голгијеву мембрану или ендоплазматски ретикулум); на овај начин, након при самосталном састављању, нуклеокапсид се приближава модификованој мембрани, почиње процес пупања и вирус бежи од тога да се делимично умота у модификовану мембрану и добије перикапсид (или омотач).
МЕХАНИЗМИ ИНТЕРАКЦИЈЕ ВИРУС-ЋЕЛИЈА: вирусна инфекција.
ПРОИЗВОДНА ИНФЕКЦИЈА: производи нове вирусе (вирусно потомство);
ОГРАНИЧАВАЈУЋЕ: вирус се размножава само када је ћелија у одређеним условима (на пример у фази С);
АБОРТИВНО: вирус се не реплицира, већ експримира само неке протеине без могућности стварања нових вириона;
ТРАЈАН: може бити хроничан - вирус се споро реплицира и ћелија ослобађа вирус током дужих периода (месеци или чак године), као у случају ХИВ -а и хроничног хепатитиса - или латентна (вирусни геном остаје тих у језгру ћелије домаћина на дуже периоде, да би се потом реактивирала да би дала продуктивну инфекцију, као у случају херпес симплекса или зостера).
ТРАНСФОРМАЦИЈА: типично за онкогене вирусе, који не убијају ћелију, већ је трансформишу у неопластичном смислу. У овим случајевима вирусни геном се интегрише у ћелијски и узима име провируса; ова промена може довести до генетске промене ћелије домаћина, која се трансформише у неопластичном смислу и, размножавајући се на неконтролисан начин, преноси аномалије на ћелије ћерке.
Вирусна инфекција може изазвати акутну болест са кратким током и некомпликованим опоравком (обично услед продуктивне инфекције, као у случају обичне прехладе), или хроничну болест.
Остали чланци о „Умножавању вируса“
- Структура и класификација вируса
- Вирус
- Болести изазване вирусима и антивирусним лековима