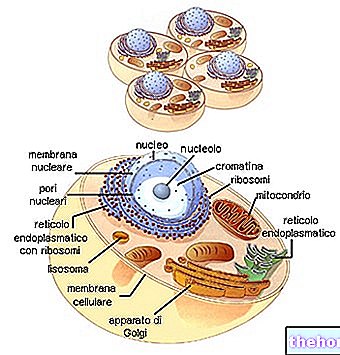
Циклични континуитет живих бића налази везе између узастопних генерација у феноменима репродукције.
Репродукција се врши на различитим нивоима еволуционе скале, у различитим гранама биљног и животињског царства, у различитим живим врстама, са толико различитих механизама који једини оправдавају читаву расправу.
Прва класификација феномена репродукције мора разликовати једноћелијске од вишећелијских организама, јер се само у првом деоба ћелије поклапа са репродукцијом.
У вишећелијским ћелијама репродукција може бити агамична или сексуална (или гамска).
Агамична репродукција, релативно ређа, заснована је на механизму митозе, тако да је варијабилност врсте поверена поновљеној појави мутација.
Постоје и различити механизми, попут стробилизације, спорулације итд., Док у биљкама налазимо регенеративне облике добро познате у пољопривреди (резнице, раслојавање итд.).
Најраспрострањенији репродуктивни механизам у вишим облицима је, међутим, полни, који одговара појави мејозе, стварању гамета и њиховом стапању у зиготи (оплодња).
Код примитивних врста гамете нису морфолошки диференциране: у овом случају говоримо о изогаметији. Међутим, увек постоје две серије гамета, идентификоване симболима (+) и (-), а оплодња се може догодити само из „сусрета гамета супротног знака: дакле, постоји биолошка разлика, која још није изражена морфолошки.
Како еволуцијска скала напредује, појављује се морфолошка и функционална диференцијација, у којој је тип женских гамета опћенито обдарен обилним резервним материјалом (деутоплазма или теле, које ће осигурати развој ембрија све док не постане метаболички неовисан) и тип мушке полне ћелије обдарене покретљивошћу да допру до женских гамета. Гаметне ћелије су увек хаплоидне и резултат су мејозе. Њиховом фузијом настаје диплоидна зигота.
Између мејозе и зиготе може проћи низ генерација хаплоидних ћелија, баш као што између зиготе и мејозе може проћи низ генерација диплоидних ћелија, при чему се различите варијације генеративних циклуса изражавају смењивањем генерација.
За врсту диплонте (са диплоидним организмом) карактеристична је гаметска мејоза: мејоза директно производи гамете које спајањем одмах реконституишу диплоидно стање. Ово је преовлађујући случај Метазоана, укључујући човека.
Гаметогенеза
Узимајући у обзир репродукцију са гаметском мејозом, каква се јавља код човека, покушајмо да разјаснимо како се мејоза уклапа у гаметогенезу (стварање гамета).
У ембрионалном развоју, мушке и женске гаметогенезе (зване сперматогенеза и оогенеза), постоји рана разлика између ћелија које су предодређене да формирају тело (соматска линија) и оних које су предвиђене за стварање гамета (заметна линија). Почетне ћелије заметне линије називају се протогони. Диференцијацијом гонада у мушком или женском смислу долази до диференцијације заметних ћелија у сперматогонију и овогонију.
Гледајући сперматогенезу, видимо да у сперматогонији постоји низ ћелијских генерација које се настављају током читавог живота. Само део тако континуирано произведене сперматогоније разликује се од нормалног митотичког циклуса, а уместо њега почиње мејотски.
Заметна ћелија у којој ће почети мејоза (редупликација, а затим прва деоба) назива се сперматоцити првог реда; његовом подјелом настају два сперматоцита другог реда, који са другим дијељењем дају укупно четири сперматида.
Можемо постепено смањити редукцију хромозомског комплета са 4н сперматоцита првог реда (након редупликације постоје четири хроматила за сваки пар хомолога) на 2н сперматоцита другог реда и на н сперматида, као што је већ виђено проучавањем мејозе, што Тако закључујемо.Сперматиде су дакле већ хаплоидне, али још нису зреле гамете.Из структуре ћелије хаплоидног типа, функционално сазревање (названо спермиохистогенеза) претвара сперматиде у сперматозоиде, односно зреле мушке гамете.
У женској гаметогенези (или оогенези) постоји неколико разлика. Пре свега, број гамета које треба припремити је много мањи. Процењује се да је око 5 Кс 105 овогона припремљено у полним жлездама женке људске врсте; од њих, само око 400 је заинтересовано за сазревање фоликула и касније избацивање, у циклусу који обично погађа само један фоликул месечно током плодног периода од око 35 година.
Различит број гамета припремљених код два пола одговара већ споменутој разлици у функцији и понашању: сперматозоиди су мали, покретни и бројни у односу на потребу тражења јајне ћелије и малу вероватноћу да ће је пронаћи; јаја су велики, инертни и малобројни, у односу на функцију гарантовања ембриону резервног материјала и заштиту коју им пружа унутрашња оплодња (природно, нарочито код спољне оплодње јајне ћелије такође морају бити бројније).
Потреба за обезбеђивањем гамета са резервним материјалом одговара присуству, у "оогенези, фазе заустављања мејозе, током које се хромозоми делимично деспирализују. Затим посматрамо такозване" пернате хромозоме ", у којима низ екстроверзија идентификује особине у којима се гени одговорни за синтезу деутоплазме депримирају.
Мањи број потребан за женске гамете такође одговара чињеници да, од четири хаплоидне ћелије произведене мејозом, само једна прима комплетан резервни материјал и постаје гамета, док остале три (полоцити или поларна тела), која садрже само хромозомски материјал, не могу да изазову зиготе и ембрионе и предодређени су за регресију.
Ђубрење
Оплодња, односно сусрет мушке и женске гамете, може се спровести на веома различите начине.У животињском царству посматрамо прелазак са спољашње оплодње (полне ћелије изложене било каквом еколошком ризику и стога нужно веома бројне у два пола) на унутрашње оплодњу, за коју је родитељска брига додатно повезана са метаболичким односом сисара између мајке и фетуса .
Оплодња, након што је дошло до сусрета између гамета супротног пола, мора се догодити гарантујући два услова: специфичност и јединственост. Односно, мора се осигурати да сперматозоид буде исте врсте као и јаје и да, кад први уђе, други неће ући.
Специфичност је осигурана биохемијским карактеристикама акросома и површине јајне ћелије. У ствари, говори се о реакцијама између «фертилисина» и «антифертилисина», са специфичношћу упоредивом са оном у сусрету између ензима и супстрата.
Јединственост оплодње загарантована је модификацијом површинске структуре јајне ћелије («кортикална реакција») која почиње одмах у тренутку прве специфичне реакције фертилисин / антифертилисин; након ове реакције, опна јајне ћелије се мења, тако да сви други сперматозоиди који дођу до ње више нису у стању да покрену специфичну реакцију оплодње.
Након оплодње, реп спермотозоана остаје ван јајне ћелије, док хромозомски материјал продире у њу. Ово, названо "мушки пронуклеус", придружује се "женском пронуклеусу" јајне ћелије формирајући тако диплоидно језгро зиготе.