Дефиниција
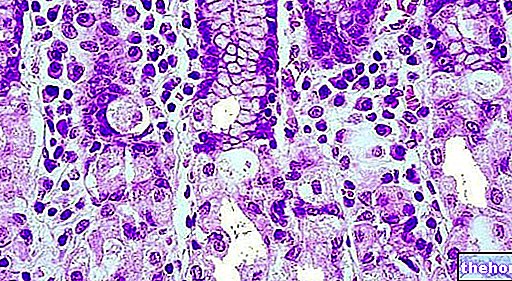
Ензими су протеини произведени у биљним и животињским ћелијама, који делују као катализатори који убрзавају биолошке реакције без промене.
Ензими делују комбинујући се са одређеном супстанцом и претварају је у другу супстанцу; класични примери дати су ензимима за варење присутним у пљувачки, желуцу, панкреасу и танком цреву, који обављају битну функцију у варењу и помажу разградњу хране на основне састојке, које тело затим може апсорбовати и искористити, прерадити другим ензимима или излучују као отпад.
Сваки ензим има посебну улогу: онај који разлаже масти, на пример, не делује на протеине или угљене хидрате. Ензими су неопходни за добробит организма. Недостатак, чак и једног ензима, може изазвати озбиљне поремећаје. Познати пример је фенилкетонурија (ПКУ), болест коју карактерише немогућност метаболизма есенцијалне аминокиселине, фенилаланин, чија акумулација може изазвати физичке деформитете и менталне болести.
Биохемијска студија
Ензими су посебни протеини који имају карактеристику да су биолошки катализатори, односно имају способност разградње енергије активације (Еатт) реакције, мењајући њен пут тако да је кинетички спор процес бржи.
Ензими повећавају кинетику термодинамички могућих реакција и, за разлику од катализатора, мање -више су специфични: стога поседују специфичност супстрата.
Ензим није укључен у стехиометрију реакције: да би се то догодило, коначно каталитичко место мора бити идентично почетном.
У каталитичком деловању скоро увек постоји спора фаза која одређује брзину процеса.
Када говоримо о ензимима, није исправно говорити о равнотежним реакцијама, уместо тога говоримо о стабилно стање (стање у којем се одређени метаболит формира и континуирано троши, одржавајући његову концентрацију скоро константном током времена). Производ реакције коју катализује један ензим је обично сам по себи реактант за наредну реакцију, катализован другим ензимом итд.
Процеси које катализују ензими обично се састоје од низа реакција.
Генеричка реакција коју катализује ензим (Е) може се сажети на следећи начин:

Е је ензим
С је подлога;
ЕС представља адукт између ензима и супстрата;
П је производ;
К је константа брзине реакције.
Генерички ензим (Е) се у комбинацији са супстратом (С) формира адукт (ЕС) са константом брзине К1; може се дисоцирати натраг у Е + С, са константом брзине К2, или, (ако "живи" довољно дуго ) може прећи у облик П са константом брзине К3.
Производ (П) се, пак, може рекомбиновати са ензимом и реформисати адукт константом брзине К4.
Када се ензим и супстрат помешају, постоји делић времена у коме се још није догодио сусрет између две врсте: то јест, постоји изузетно кратак временски интервал (који зависи од реакције) у коме ензим и супстрат имају још није испуњен; након овог периода ензим и супстрат долазе у контакт у све већим количинама и формира се ЕС адукт. Након тога, ензим делује на супстрат и производ се ослобађа. Тада се може рећи да је ц "почетни временски интервал у којем се концентрација ЕС адукта не може дефинисати; након овог периода, претпоставља се да је стабилно стање је утврђено, односно брзина процеса који доводе до добијања адукта једнака је брзини процеса који доводе до уништења адукта.
Мицхаелис-Ментенова константа (КМ) је константа равнотеже (упућује на прву горе описану равнотежу); може се рећи, са добром апроксимацијом (јер треба узети у обзир и К3), да је КМ представљен односом између кинетичких константи К2 и К1 (односи се на уништавање и формирање адукта ЕС у првој горе описаној равнотежи) .
Кроз Мицхаелис-Ментенову константу имамо "индикацију афинитета између ензима и супстрата: ако је КМ мала ц" је "висок афинитет између ензима и супстрата, онда је ЕС адукт стабилан.
Ензими подлежу регулацији (или модулацији).
У прошлости се углавном говорило о негативној модулацији, односно инхибицији каталитичких способности ензима, али може постојати и позитивна модулација, односно постоје врсте способне да повећају каталитичке способности ензима.
Постоје 4 врсте инхибиција (добијене из апроксимација направљених на моделу ради усклађивања експерименталних података са математичким једначинама):
- конкурентна инхибиција
- неконкурентна инхибиција
- неконкурентна инхибиција
- конкурентна инхибиција
Говоримо о компетитивној инхибицији када је молекул (инхибитор) способан да се такмичи са супстратом. Због структурне сличности, инхибитор може реаговати уместо супстрата; отуда и терминологија "компетитивна инхибиција". Вероватноћа да се ензим веже за инхибитор или супстрат зависи од концентрације оба и њиховог афинитета са ензимом; брзина реакције стога зависи од ових фактора.
Да би се постигла иста брзина реакције као и без присуства инхибитора, потребно је имати већу концентрацију супстрата.
Експериментално је показано да се, у присуству инхибитора, Мицхаелис-Ментенова константа повећава.
Што се тиче, уместо тога, „неконкурентна инхибиција, интеракција између молекула који би требало да функционише као модулатор (позитиван или негативан инхибитор) и„ ензима, одвија се на месту које је различито од оног на коме је интеракција јавља се између ензима и супстрата; стога говоримо о алостеричној модулацији (из грчког аллостерос → други сајт).
Ако се инхибитор веже за ензим, он може изазвати промену структуре ензима и, сходно томе, може смањити ефикасност којом се супстрат везује за ензим.
У овој врсти процеса, Мицхаелис-Ментенова константа остаје константна јер ова вредност зависи од равнотеже између ензима и супстрата, па се чак и у присуству инхибитора те равнотеже не мењају.
Феномен неконкурентне инхибиције је редак; типичан некомпетитивни инхибитор је супстанца која се реверзибилно везује за ЕС адукт узрокујући ЕСИ:

Инхибиција вишка супстрата понекад може бити неконкурентна, јер се то дешава када се други молекул супстрата веже за ЕС комплекс, стварајући ЕСС комплекс.
Акомпетитивни инхибитор, с друге стране, може се везати само за супстрат ензимски адукт као у претходном случају: везивање супстрата за слободни ензим индукује конформациону модификацију која чини место доступним за инхибитор.
Константа Мицхаелис Ментен опада са повећањем концентрације инхибитора: очигледно, дакле, расте афинитет ензима за супстрат.
Серин протеаза
Они су породица ензима којима припадају кимотрипсин и трипсин.
Кимотрипсин је протеолитички и хидролитички ензим који сече десно од хидрофобних и ароматичних аминокиселина.
Производ гена који кодира химотрипсин није активан (активира се наредбом); неактивни облик химотрипсина представљен је полипептидним ланцем од 245 аминокиселина. Химотрипсин има глобуларни облик због пет дисулфидних мостова и других мањих интеракција (електростатичке, Ван дер Ваалсове силе, водоничне везе итд.).
Кимотрипсин производе химозне ћелије панкреаса, где се налази у посебним мембранама и избацује кроз канал панкреаса у црева, у време варења хране: кимотрипсин је у ствари дигестивни ензим. Протеини и хранљиве материје које уносимо исхраном подвргавају се варењу како би се свели на мање ланце и апсорбовали и претворили у енергију (нпр. Амилазе и протеазе разграђују хранљиве материје у глукозу и аминокиселине које до крвних судова доспевају у ћелије стижу до порталне вене и одатле се транспортују у јетру где се подвргавају даљем третману).
Ензими се производе у неактивном облику и активирају се тек када стигну на „место на коме морају да раде“; када се њихова радња заврши, деактивирају се. Ензим, једном деактивиран, не може се поново активирати: да би имао „даље каталитичко дејство, мора га заменити“ другим молекулом ензима. Ако би се химитрипсин већ произвео у активном облику у панкреасу, он би напао потоњи: панкреатитис су патологије узроковане пробавним ензимима који су већ активирани у панкреасу (а не на потребним местима); неки од њих ако се не лече на време, довести до смрти.
У кимотрипсину и у свим серинским протеазама, каталитичко деловање је последица постојања алкохолног аниона (-ЦХ2О-) у бочном ланцу серина.
Серинске протеазе узимају ово име управо зато што је њихово каталитичко деловање последица серина.
Када сви ензими изврше своју акцију, пре него што поново могу да оперишу супстрат, морају се обновити водом; "ослобађање" серина водом је најспорија фаза процеса, а то је та фаза који одређује брзину катализе.
Каталитичко дејство се одвија у две фазе:
- формирање аниона са каталитичким својствима (ањонски алкохолат) и накнадни нуклеофилни напад на карбонил угљеник (Ц = О) са цепањем пептидне везе и формирањем естара;
- водени напад са обнављањем катализатора (способан да поново изврши своје каталитичко дејство).
Различити ензими који припадају породици серинских протеаза могу се састојати од различитих аминокиселина, али за све њих каталитичко место представља алкохолни анион бочног ланца серина.
Потпородица серинских протеаза је она ензима укључених у коагулацију (која се састоји у трансформацији протеина, из њиховог неактивног облика у "други облик који је активан"). Ови ензими осигуравају да је коагулација што је могуће ефикаснија и ограничена у простор и време (коагулација се мора појавити брзо и мора се десити само у близини повређеног подручја) .Ензими укључени у коагулацију активирају се каскадно (активирањем једног ензима добијају се милијарде ензима: сваки активирани ензим , заузврат активира многе друге ензиме).
Тромбоза је патологија услед неисправног деловања ензима коагулације: узрокована је активацијом, без потребе (јер нема повреде), ензима који се користе у коагулацији.
Постоје модулирајући (регулаторни) ензими и инхибиторни ензими за друге ензиме: у интеракцији са последњим регулишу или инхибирају њихову активност; чак и производ ензима може бити инхибитор ензима.Има и ензима који делују што је супстрат већи.
Лизоцим
Луиги Пастеур је кихањем на петријеву здјелу открио да у слузи постоји ензим који може убити бактерије: лизоцим; са грчког: лисо = које величине; зимо = ензим.
Лизозим је способан да разбије ћелијски зид бактерија. Бактеријама и једноћелијским организмима уопште требају механички отпорне структуре које ограничавају њихов облик; унутар бактерија постоји веома висок осмотски притисак па привлаче воду. Плазма мембрана би експлодирала да нема ћелијског зида који се противи уласку воде и ограничава волумен бактерије.
Ћелијски зид се састоји од полисахаридног ланца у коме се смењују молекули Н-ацетил-глукозамина (НАГ) и молекули Н-ацетил-мураминске киселине (НАМ); веза између НАГ и НАМ се прекида хидролизом.Карбоксилна група НАМ, у ћелијском зиду, ангажована је у пептидној вези са амино киселином.
Између различитих ланаца формирају се мостови који се састоје од псеудо-пептидних веза: гранање је последица молекула лизина; структура у целини је веома разграната и то јој даје високу стабилност.
Лизозим је антибиотик (убија бактерије): делује тако што прави пукотину у бактеријском зиду; када се ова структура (која је механички отпорна) сломи, бактерија извлачи воду све док не пукне. Лизозим успева да разбије β-1,4 глукозидну везу између НАМ и НАГ.
Каталитичко место лизозима представљено је жлебом који пролази дуж ензима у који је уметнут полисахаридни ланац: шест глукозидних прстенова ланца смештено је у жлеб.
У положају три утора ц "налази се пригушивач: у овом положају може се поставити само један НАГ, јер НАМ, који је већих димензија, не може ући. Стварно каталитичко место је између положаја четири и пет: будући да постоји НАГ на позицији три, рез ће се одвијати између НАМ и НАГ (а не обрнуто); рез је, према томе, специфичан.
Оптимални пХ за деловање лизозима је пет. На каталитичком месту ензима, односно између положаја четири и пет, налазе се бочни ланци аспарагинске и глутаминске киселине.
Степен хомологије: мери сродство (тј. Сличност) између протеинских структура.
Постоји снажна веза између лизозима и лактоза-синтазе.
Лактоза синтетаза синтетише лактозу (која је главни млечни шећер): лактоза је галактозил глукозид у коме је ц "β-1,4 глукозидна веза између галактозе и глукозе.
Стога, лактоза синтетаза катализује супротну реакцију од оне коју катализује лизозим (који уместо тога цепа β-1,4 глукозидну везу)
Лактоза синтетаза је димер, односно састоји се од два протеинска ланца, од којих један има каталитичка својства и упоредив је са лизозимом, а други је регулаторна подјединица.
Током трудноће, ћелије млечне жлезде синтетишу гликопротеине дејством галатозил-трансферазе (има „хомологију секвенце од 40% са лизозимом): овај ензим је у стању да пренесе галактозил групу из високоенергетске структуре у структура гликопротеина.Током трудноће индукује се експресија гена који кодира галактозил-трансферазу (постоји и експресија других гена који такође дају друге производе): долази до повећања величине дојке јер се активира млечна жлезда (претходно неактивна) која мора да производи млеко. Током порођаја се производи α-лакталбумин који је регулаторни протеин: у стању је да регулише каталитички капацитет галактозил-трансферазе (дискриминацијом супстрата). Галактозил-трансфераза модификована α-лакталалбумином је у стању да пренесе галактозил на молекул глукозе: формира β-1,4 гликозидну везу и даје лактозу (лактоза синтетаза).
Дакле, галактоза трансфераза припрема млечну жлезду пре порођаја и производи млеко након порођаја.
За производњу гликопротеина, галактозил трансфераза се везује за галактозил и НАГ; током порођаја лактални албумин се веже за галактозилтрансферазу чинећи да овај препозна глукозу и више не НАГ да даје лактозу.




-effetti-sulla-salute.jpg)