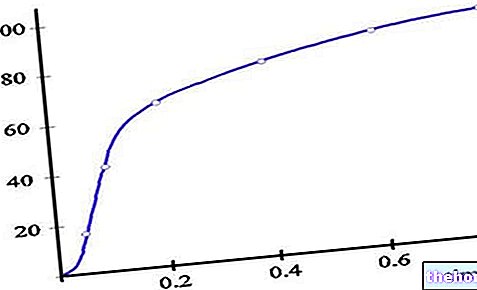
Аминокиселине и протеини су посредници преласка из света минерала у живу материју.
Као што им назив говори, аминокиселине су бифункционалне органске супстанце: састоје се од амино функције (-НХ2) и карбоксилне функције (-ЦООХ); могу бити α, β, γ итд., у зависности од положаја који амино група заузима у односу на карбокси групу:



Све биолошки важне аминокиселине су α-аминокиселине.
Протеинске структуре се састоје од двадесет аминокиселина.
Као што се може видети из горе приказаних генеричких структура, све аминокиселине имају заједнички део и другачији део који их карактерише (генерално представљене са Р).
Од двадесет аминокиселина, деветнаест је оптички активно (одбија равнину поларизоване светлости).
Већина аминокиселина има само једну амино групу и један карбоксил па се зато и зову неутралне аминокиселине; зову се они који имају додатни карбоксил киселе аминокиселине док су они са екстра амино групом базичне аминокиселине.
Аминокиселине су кристалне чврсте материје и имају добру растворљивост у води.
Недостатак неких аминокиселина у исхрани изазива озбиљне промене у развоју; у ствари, људски организам није у стању да синтетише неке аминокиселине које се тачно називају есенцијалне (морају се увести исхраном), док може сам произвести само неке аминокиселине (оне које нису есенцијалне).
Једна од болести због недостатка есенцијалних аминокиселина је она позната под именом квасхиоркор (реч која потиче из афричког дијалекта што у преводу значи „прва и друга“); ова болест погађа прворођенче, али након рођења другог детета јер првом детету недостаје мајчино млеко које садржи правилну количину протеина. Ова болест је, дакле, широко распрострањена међу неухрањеном популацијом и укључује дијареју, недостатак апетита који доводи до прогресивног слабљења организма.
Као што је већ поменуто, природне аминокиселине, са изузетком глицина (то је а-аминокиселина са водоником уместо Р групе и најмања је од двадесет), поседују оптичку активност због присуства најмање једне асиметричне угљеник. У природним аминокиселинама апсолутна конфигурација асиметричног угљеника на који су повезане само амино и карбокси групе припада Л серији;
Д-аминокиселине никада не постају део структуре протеина.
Сећамо се тога:
ДНК ---- транскрипција → м-РНК ---- превод → протеин
Транскрипција се може кодирати као Л-аминокиселине; Д-аминокиселине могу бити садржане у не-протеинским структурама (нпр. У зиду бактерије: у бактеријама не постоје "генетске информације о томе да Д-аминокиселине имају заштитну улогу, међутим, постоје" генетске информације за ензиме који се баве зидом бактеријске облоге).
Вратимо се аминокиселинама: различита структура Р групе дефинише индивидуалне карактеристике сваке аминокиселине и даје посебан допринос карактеристикама протеина.
Због тога се сматрало да се аминокиселине деле на основу природе Р групе:
Поларне, али ненаелектрисане аминокиселине:
Глицин (Р = Х-)
Серин (Р = ХО-ЦХ2-)
Треонинбитан

Треонин има два центра симетрије: само 2С, 3Р треонин постоји у природи.
Треонин је есенцијална аминокиселина (не треба мешати са неопходном: све аминокиселине су есенцијалне), па се мора узимати са исхраном, односно конзумирањем хране која га садржи јер, као што је већ поменуто, генетско наслеђе није присутно у ћелије људских бића. способне да производе ову аминокиселину (ово наслеђе је присутно у многим биљкама и бољим).
Хидроксилна група серина и треонина може се естерификовати фосфорилном групом (добијајући фосфосерин и фосфотреонин), овај процес се назива фосфорилација; фосфорилација се у природи користи за превођење сигнала између унутрашњости и спољашњости ћелије.
Цистеин (Р = ХС-ЦХ2-)
Сулфхидрил цистеина лакше се може протонирати од хидроксила серина: сумпор и кисеоник су шесте групе, али се сумпор лакше оксидује јер има веће димензије.
Тирозин [Р = ХО- (Ц6Х4) -ЦХ2-]
Н.Б.
(Ц6Х4) = ди-супституисани бензенски прстен
Као и код серина и треонина, хидроксил се може естерификовати (фосфорилисати).Аспарагин (Р = НХ2-ЦО-ЦХ2-)
Глутамин (Р = НХ2-ЦО-ЦХ2-ЦХ2-)
Неполарне аминокиселине
имају хидрофобне бочне групе; унутар ове класе разликујемо:
Алифатика:
Аланин (Р = ЦХ3-)
Валин (Р = (ЦХ3) 2-ЦХ-) неопходан
Леуцин (Р = (ЦХ3) 2-ЦХ-ЦХ2-) неопходан
Изолеуцин (Р =
) битан
Метионин (Р = ЦХ3-С-ЦХ2-ЦХ2-) неопходан

Ћелијске мембране се састоје од липидног двослоја са протеинима усидреним захваљујући њиховом хидрофобном карактеру, па садрже аланин, валин, изолеуцин и леуцин. С друге стране, метионин је аминокиселина која је скоро увек присутна у малим количинама (око 1%).
Пролине

Ароматично:
Фенилаланин (Р = Пх-ЦХ2-) Пх = фенил: есенцијални моносупституисани бензен
Триптофан (Р =
битан

Ове две аминокиселине, будући да су ароматичне, апсорбују близу ултраљубичастог зрачења (око 300 нм); стога је могуће искористити технику УВ спектрофотометрије за одређивање концентрације познатог протеина који садржи ове аминокиселине.
Напуњене аминокиселине
Заузврат се деле на:
Киселе аминокиселине (које имају поларне остатке са негативним набојем при пХ 7) су такве зато што могу да дају позитиван Х + набој:
Аспарагинска киселина
Глутаминска киселина (Р =
)
Ове аминокиселине потичу од аспарагина и глутамина; сва четири постоје у природи и то значи да је ц "специфична информација за сваког од њих, то јест ц" је базна тројка у ДНК која кодира сваку од њих.
Базне аминокиселине (које имају поларне остатке са позитивним набојем при пХ 7) су такве зато што су способне да прихвате позитиван Х + набој:
Лизин (Р =
) битан
Аргинин (Р =
)
Хистидин (Р =
)





Постоје протеини у којима у бочним ланцима постоје деривати аминокиселина: на пример, може бити присутан фосфосерин (не постоје генетске информације које кодирају фосфосерин, само оне за серин); фосфосерин је модификација пост-транслацијски: након што је дошло до синтезе протеина
ДНК ---- транскрипција → м-РНК ---- превод → протеин
такве пост-транслационе модификације могу се појавити на бочним ланцима протеина.
Такође погледајте: Протеини, поглед на хемију